
Трансмембранный электрохимический потенциал как промежуточная форма энергии при окислительном фосфорилировании
На втором этапе синтезируется АТФ.
Синтез АТФ.Фермент КГ-транслоцирующая АТФ синтаза состоит из двух крупных белковых комплексов, каждый из которых построен из многих субъединиц. В структуре этого фермента выделяют протонный канал (Fo) и каталитическую часть фермента (Fi) содержащую три активных центра. В каждый из активных-центров поступают молекулы АДФ и неорганического фосфата (Pi) Протонный канал пронизывает внутреннюю митохондриальную мембрану, а каталитическая часть фермента представляет собой крупное грибовидное образование, обращенное в сторону матрикса. Ультразвуковая обработка интактных (неповрежденных) митохондрий вызывает остановку синтеза АТФ, поскольку каталитическая часть фермента Fi отделяется от протонного канала. При этом поступление потока протонов из межмембранного пространства в митохондриальныи матрикс не нарушается.
Под действием потока протонов по каналу каталитическая часть АТФ синтазы (Fo) вращается вокруг устья канала. Синтез АТФ проходит три стадии:
1) поступление АДФ и неорганического фосфата (Р;) в каталитический центр,
2) поворот на 120° и сжимающая деформация каталитического центра, сближаю
щая отталкивающиеся группы остатков фосфорной кислоты; на этой стадии про
исходит образование АТФ и молекулы воды,
3) поворот на 120° и освобождение из активного центра молекулы АТФ.
Затем следует поворот на 120° АТФ синтазы и в освободившийся каталитический центр поступают новые молекулы АДФ и Pi (неорганического фосфата)
Структурная организация дыхательной цепи




![]()
![]()
Все комплексы дыхательной цепи интегрированы во внутренней мембране митохондрий (рис. 21). Протон генерирующими являются комплексы I, III и IV.
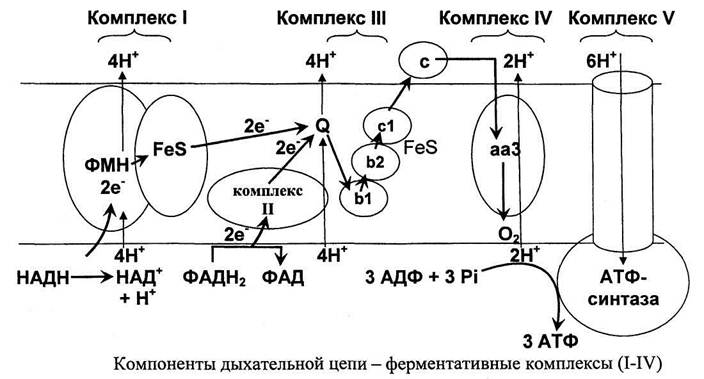
Образующиеся при окислении сукцината, ацетил-КоА и других субстратов электроны переносятся на убихинон (Q) комплексом II через связанный с ферментом ФАДНг или через электрон переносящий флавопротеин (EFT). Прохождение пары электронов через комплекс II сопровождается переносом 4-х протонов из матрик-са.
Трансмембранный электрохимический потенциал как промежуточная форма энергии при окислительном фосфорилировании
На втором этапе синтезируется АТФ.
Синтез АТФ.Фермент КГ-транслоцирующая АТФ синтаза состоит из двух крупных белковых комплексов, каждый из которых построен из многих субъединиц. В структуре этого фермента выделяют протонный канал (Fo) и каталитическую часть фермента (Fi) содержащую три активных центра. В каждый из активных-центров поступают молекулы АДФ и неорганического фосфата (Pi) Протонный канал пронизывает внутреннюю митохондриальную мембрану, а каталитическая часть фермента представляет собой крупное грибовидное образование, обращенное в сторону матрикса. Ультразвуковая обработка интактных (неповрежденных) митохондрий вызывает остановку синтеза АТФ, поскольку каталитическая часть фермента Fi отделяется от протонного канала. При этом поступление потока протонов из межмембранного пространства в митохондриальныи матрикс не нарушается.
Под действием потока протонов по каналу каталитическая часть АТФ синтазы (Fo) вращается вокруг устья канала. Синтез АТФ проходит три стадии:
1) поступление АДФ и неорганического фосфата (Р;) в каталитический центр,
2) поворот на 120° и сжимающая деформация каталитического центра, сближаю
щая отталкивающиеся группы остатков фосфорной кислоты; на этой стадии про
исходит образование АТФ и молекулы воды,
3) поворот на 120° и освобождение из активного центра молекулы АТФ.
Затем следует поворот на 120° АТФ синтазы и в освободившийся каталитический центр поступают новые молекулы АДФ и Pi (неорганического фосфата)
39.Окислительное фосфорилирование, коэффициент р/о. Строение митохондрий и структурная организация дыхательной цепи. Трансмембранный электрохимический потенциал.
Синтез АТФ из АДФ и Н3РО4за счёт энергии переноса электронов по ЦПЭ называютокислительным фосфорилированием.
Протонный градиент и электрохимический потенциал Перенос электронов по дыхательной цепи от NADH к кислороду сопровождается выкачиванием протонов из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство. На эту работу затрачивается часть энергии электронов, переносимых по ЦПЭ.
Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμH + заставляет протоны двигаться по каналу АТФ-синтазы в матрикс. Параллельно под действием ΔμH + происходят конформационные изменения в парах α, β-субъединиц белка F1, в результате чего из АДФ и неорганического фосфата образуется АТФ. Электрохимический потенциал, генерируемый в каждом из 3 пунктов сопряжения в ЦПЭ, используют для синтеза одной молекулы АТФ.
Каждая митохондрия окружена оболочкой, состоящей из двухмембран; между ними — межмембранное пространство. Отграниченное внутренней мембраной пространство называется матриксом. В матриксе содержатся большая частьферментов, участвующих вцикле Кребса, протекаетокислениежирных кислот, располагаются митохондриальныеДНК,РНКирибосомы. Внутренняя мембрана образует многочисленные гребневидные складки — кристы, существенно увеличивающие площадь ее поверхности. Наружная мембрана митохондрий имеет маленькие отверстия, образованные специальными белками, через которые могут проникать небольшиемолекулыиионы. Внутренняя мембрана таких отверстий не имеет; на ней, на стороне, обращенной к матриксу, располагаются особые молекулыАТФ-синтазы, состоящие из головки, ножки и основания. При прохождении через нихпротоновпроисходит синтез АТФ. В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи. Наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
4.8.10.Окислительное фосфорилирование, коэффициент р/о. Строение митохондрий и структурная организация дыхательной цепи. Трансмембранный электрохимический потенциал.
Синтез АТФ из АДФ и Н3РО4за счёт энергии переноса электронов по ЦПЭ называютокислительным фосфорилированием.
Протонный градиент и электрохимический потенциал Перенос электронов по дыхательной цепи от NADH к кислороду сопровождается выкачиванием протонов из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство. На эту работу затрачивается часть энергии электронов, переносимых по ЦПЭ.
Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμH + заставляет протоны двигаться по каналу АТФ-синтазы в матрикс. Параллельно под действием ΔμH + происходят конформационные изменения в парах α, β-субъединиц белка F1, в результате чего из АДФ и неорганического фосфата образуется АТФ. Электрохимический потенциал, генерируемый в каждом из 3 пунктов сопряжения в ЦПЭ, используют для синтеза одной молекулы АТФ.
Каждая митохондрия окружена оболочкой, состоящей из двухмембран; между ними — межмембранное пространство. Отграниченное внутренней мембраной пространство называется матриксом. В матриксе содержатся большая частьферментов, участвующих вцикле Кребса, протекаетокислениежирных кислот, располагаются митохондриальныеДНК,РНКирибосомы. Внутренняя мембрана образует многочисленные гребневидные складки — кристы, существенно увеличивающие площадь ее поверхности. Наружная мембрана митохондрий имеет маленькие отверстия, образованные специальными белками, через которые могут проникать небольшиемолекулыиионы. Внутренняя мембрана таких отверстий не имеет; на ней, на стороне, обращенной к матриксу, располагаются особые молекулыАТФ-синтазы, состоящие из головки, ножки и основания. При прохождении через нихпротоновпроисходит синтез АТФ. В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи. Наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
НАД-зависимые и флавиновые дегидрогеназы, убихинон-дегидрогеназа, цитохромы в, с, с1, а1 и а3 как компоненты дыхательной цепи.
Эти коферменты входят в состав активных центров дегидрогеназ, но могут обратимо диссоциировать из комплекса с апоферментами и включаются в состав фермента в ходе реакции. Субстраты NAD- и NADP-зависимых дегидрогеназ находятся в матриксе митохондрий и в цитозоле. Рабочей частью никотинамидных коферментов служит никотинамид.
NADPH + NAD + ↔ NADP + + NADH.

Флавиновые дегидрогеназы содержат в качестве коферментов FAD или FMN. Эти коферменты образуются в организме человека из витамина В2. Флавиновые коферменты прочно связаны с апоферментами. Рабочей частью FAD и FMN служит изоаллоксазиновая сопряжённая циклическая система.
FAD служит акцептором электронов от многих субстратов в реакциях типа:
Цепь переноса электронов от NADH и FADH2 на кислород Перенос электронов от NADH к О2 включает ряд переносчиков, которые локализованы во внутренней мембране митохондрий. За исключением убихинона и цитохрома С, это сложные белковые комплексы.
NADH + Н + + Е (FMN) → NAD + + Е (FMNH2)
С FMNH2 электроны переносятся затем на ряд железо-серных белков (FeS), играющих роль второй простетической группы в молекуле NADH-дегидрогеназы. Атомы железа в этих белках (негемовое железо) собраны в несколько групп, так называемых железо-серных центров. FeS-центры входят в состав многих белков (флавопротеинов, цитохромов), участвующих в окислительно-восстановительных реакциях. Известны 3 типа FeS-центров (FeS, Fe2S2, Fe4S4), в которых атом железа связан с атомом серы остатков цистеина или неорганической серы. NADH-дегидрогеназа содержит несколько центров типа Fe2S2 и Fe4S4 Атомы железа в таких центрах могут принимать и отдавать электроны поочерёдно, переходя в ферро- (Fe2+) и ферри- (Fe3+) состояния. От железо-серных центров электроны переносятся на кофермент Q (убихинон).

различием боковых цепей в структуре тема;
различием в структуре полипептидных цепей;
различием в способе связи полипептидных цепей с гемом.
В зависимости от способности поглощать свет в определённой части спектра все цитохромы делят на группы а, b, с. Внутри каждой группы отдельные виды с уникальными спектральными свойствами обозначают цифровыми индексами (b, b1, b2 и т.д.).
Структурные особенности разных видов цитохромов определяют различие в их окислительно-восстановительных потенциалах. В ЦПЭ участвуют 5 типов цитохромов (а, а3, b, с, с1). За исключением цитохрома с, все цитохромы находятся во внутренней мембране митохондрий в виде сложных белковых комплексов.
QН2-дегидрогеназа (коэнзим Q-цитохром с-ре-уктаза, комплекс III) состоит из 2 типов цитохромов (b1 и b2) и цитохрома с1. QН2-дегидрогеназа переносит электроны от убихинола на цитохром с. Внутри комплекса III электроны передаются от цитохромов b на FeS-центры, на цитохром с1, а затем на цитохром с. Группы тема, подобно FeS-центрам, переносят только по одному электрону. Таким образом, от молекулы QH2 2 электрона переносятся на 2 молекулы цитохрома b. В качестве промежуточного продукта в этих реакциях переноса электронов возможно образование свободного радикала се-михинона. В цитохромах типа b гем не связан ковалентно с белком, а в цитохромах с1 и с он присоединяется к белку при помощи тиоэфирных связей. Эти связи образуются путём присоединения 2 цистеиновых остатков к винильным группам гема.
Cu + ↔ Cu 2+ + e,
Fe 2+ ↔ Fe 3+ + e.
Комплекс цитохромов а-а3 непосредственно реагирует с молекулярным кислородом.

45. Строение митохондрий и структурная организация дыхательной цепи. Трансмембранный электрохимический потенциал как промежуточная форма энергии при окислительном фосфорилировании. Каждая митохондрия окружена оболочкой, состоящей из двух мембран; между ними — межмембранное пространство. Отграниченное внутренней мембраной пространство называется матриксом. В матриксе содержатся большая часть ферментов, участвующих в цикле Кребса, протекает окисление жирных кислот, располагаются митохондриальные ДНК,РНК и рибосомы. Внутренняя мембрана образует многочисленные гребневидные складки — кристы, существенно увеличивающие площадь ее поверхности. Наружная мембрана митохондрий имеет маленькие отверстия, образованные специальными белками, через которые могут проникать небольшие молекулы и ионы. Внутренняя мембрана таких отверстий не имеет; на ней, на стороне, обращенной к матриксу, располагаются особые молекулы АТФ-синтазы, состоящие из головки, ножки и основания. При прохождении через них протонов происходит синтез АТФ. В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи. Наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
Протонный градиент и электрохимический потенциалПеренос электронов по дыхательной цепи от NADH к кислороду сопровождается выкачиванием протонов из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство. На эту работу затрачивается часть энергии электронов, переносимых по ЦПЭ.